
欢迎光临诺贝苏(大连)营养食品有限公司企业网站!
消费者咨询热线:400-1089-520
李雪,张捷*,周鹏 (江南大学食品学院,江苏 无锡 214122)
摘 要:免疫球蛋白作为儿童免疫系统的核心武器,在儿童免疫系统发育成熟前提供保护作用,从而降低感染风险。这种活性成分作为牛乳中免疫活性的重要组成部分在初乳中含量较高。它们是母亲将被动免疫转移给后代时发生免疫联系的核心,其转移机制因哺 乳动物物种而异。无论是针对同源物种还是跨物种,免疫球蛋白的应用机制受到免疫球蛋白的类别分型、分泌过程的分子机制以及 摄入者年龄阶段等因素的影响而有所不同。牛乳已成为有益于人类的免疫产品的重要潜在来源。免疫球蛋白在牛乳加工过程中或在 胃肠道消化过程中的稳定性是评估牛乳免疫球蛋白价值的另一个考虑因素,因此,文章重点总结了牛乳中免疫球蛋白含量及其发挥 免疫作用的机制和影响,并强调了免疫球蛋白在胃肠道中的抗消化稳定性,以此为儿童免疫发育提供独特的营养需求,并提高机体免疫防御功能。
关键词:免疫球蛋白;初乳;跨物种应用;被动转移;消化特性
中图分类号:TS252.1 文献标识码:B doi:10.19827/j.issn1001-2230.2025.11.003 文章编号:1001-2230(2025)11-0022-09
LI Xue, ZHANG Jie*, ZHOU Peng (College of Food Science and Technology, Jiangnan University, Wuxi 214122, China)
Abstract:Immunoglobulins serve as the core weaponry of the pediatric immune system, providing protective effects until the immune system matures, thereby reducing the risk of infections. As a critical component of the immune activity in bovine milk, these active ingredients are present at higher concentrations in colostrum. They are central to the immune linkage that occurs during the transfer of passive immunity from the mother to the offspring, with the transfer mechanism varying among mammalian species. Bovine milk emerges as a significant potential source of immune-beneficial products for humans. The mechanism of immunoglobulin application, whether targeting homologous species or used across species, is influenced by factors such as the immunoglobulin class typing, the molecular mechanisms underlying the secretion process, and the age stage of the consumer. Simultaneously, the stability of immunoglobulins during bovine milk process or in the gastrointestinal digestion process is another consideration of assessing the value of bovine milk immunoglobulins. Therefore, this article focuses on summarizing the content of immunoglobulin in bovine milk, the mechanisms and influence through which the immunoglobulin exerts its immune effects. It also emphasizes the anti-digestive stability of immunoglobulins in the gastrointestinal tract, thus addressing the unique nutritional needs for pediatric immune development and enhancing the body’s immune defense functions.
Key words:Immunoglobulins; Colostrum; Cross-species applications; Passive transfer; Digestive stability
0 引 言
初乳中含有丰富的营养物质活性成分(尤其是免疫球蛋白和抗体)以及抗菌成分(乳铁蛋白、 抗菌蛋白、溶菌酶和乳过氧化物酶等),可以为新生儿提供被动免疫[1-2]。其中,免疫球蛋白是由哺乳动物响应抗原或免疫原性刺激(如病毒或细菌 等)合成的抗体,从而能够抵抗微生物入侵,提供免疫防御功能。免疫球蛋白是由浆细胞分泌的具 有抗菌功能的球状蛋白家族,通过特异性抗原结合能力介导中和,调理及补体激活等发挥免疫效应。哺乳动物乳汁中含有丰富的免疫球蛋白,以特异性免疫球蛋白(Immunoglobulin A,IgA)、免疫球蛋白(Immunoglobulin G,IgG)和免疫球蛋白 (Immunoglobulin M,IgM)形式为新生儿在免疫发育过程中提供被动免疫力,使其免受周围微生物和病原体的侵害[3]。母体主要通过2种方式将免疫球蛋白传递给新生儿:(1)子宫内传递,母体的免疫球蛋白通过胎盘进入胎儿的发育系统中;(2)初乳传递, 免疫球蛋白由乳腺分泌到初乳中,经新生儿摄取后进入胃肠道(Gastrointestinal,GI)[4-5]。由于健康牛和水牛在分娩后初乳中免疫球蛋白含量远大于牛犊的需求,其初乳中的免疫球蛋白作为主要免疫成分有潜力应用于婴幼儿配方奶粉、特殊膳食补充剂和创 新功能性食品中。
IgG 通过新生儿Fc受 体(Neonatal Fcreceptor, FcRn)介导的转运过程进入黏膜分泌物中,相对于IgA 通过聚合物免疫球蛋白受体的单向转运,IgG通过 FcRn 的转运在极化上皮细胞中是双向的。FcRn 不仅能够将抗原特异性 IgG 递送和分泌到黏膜表面,还负责将抗原-IgG复合物运输和回收到固有层。 IgG 的这种双向转移作用体现出其具有蛋白质的双重治疗策略[6-7]。其中,一方面,将 FcRn 修饰为 Fc 融合蛋白,促进某些药物穿过黏膜屏障的递送并增加其循环半衰期,另一方面,设计特异性靶向新生儿的 FcRn 抗体,可以抑制免疫的过度反应,从而用于治疗 IgG 和抗原抗体复合物介导的自身免疫性疾病[8]。随着免疫球蛋白以营养补充剂的形式被引入奶粉市场, 了解免疫球蛋白对儿童免疫发育和抵抗胃肠感染的 机制至关重要。本文总结了免疫球蛋白在初乳中的作用以及最新研究进展及治疗策略,有助于拓展免疫球蛋白在婴幼儿配方奶粉中的应用潜力。
1 乳中免疫球蛋白概述
1.1 乳中免疫球蛋白含量
根据免疫球蛋白的转移途径,可大概将哺乳动物分为3种类型,人类和兔子、反刍动物及部分家畜(如 牛、羊、马和猪)。其中,人类和兔子能够通过胎盘传递免疫球蛋白,研究发现新生儿血清中免疫球蛋白的含量和浓度与母体相似。其他则是从初乳中直接获得免疫球蛋白。新生儿在出生时自身免疫系统尚未完全发育,本身并不含有抗体,其获得的早期免疫保 护主要依赖于母体初乳中的免疫球蛋白。 在人类的初乳和成熟乳中测定发现,初乳中IgG的平均浓度为0.4mg/mL,IgM为0.3mg/mL,而在成熟乳中分别为0.04mg/mL和0.03mg/mL。牛初乳中免疫球蛋白占总蛋白含量的70%~80%,而在成熟乳中,免疫球蛋白仅占总蛋白的1%~2%。牛初乳中超过90%的免疫球蛋白是IgG,首次挤奶后的平均浓度约为60mg/mL(范围为15~180mg/mL)[9]。牛初乳中IgA含量在1~6mg/mL 之间,IgM含量在3~5mg/mL 之间,而成熟乳中分别为0.13mg/mL和0.04mg/mL。牛初乳中IgG的主要亚类是IgG1,约占所有免疫球蛋白的50%~80%。人类初乳和成熟乳与牛初乳和成熟乳中的免疫球蛋白浓度见表1。基于此,一些基于初乳的产品也被作为膳食补充剂来销售。免疫乳制品已被证明可有效预防各种GI病原体引起的感染[10]。化学防腐剂不能很好的保存初乳,而冷冻干燥和喷雾干燥在控制条件下可以保留更多的活性免疫球蛋白。
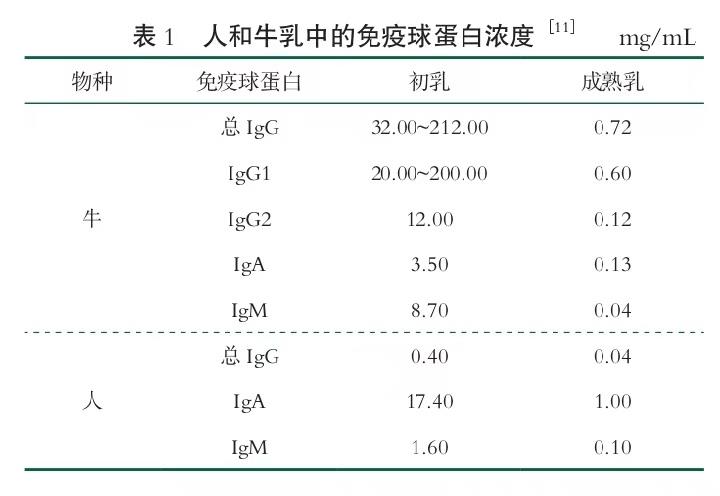
1.2 乳中免疫球蛋白结构和功能
免疫球蛋白是具有保护性的一种蛋白质化合物。 牛乳和血液的免疫球蛋白具有生物同源性。哺乳动物分泌物中有5种免疫球蛋白,根据结构和生物活性分为IgG、IgA、IgM、免疫球蛋白D(Immunoglobulin D,IgD)和免疫球蛋白E(Immunoglobulin E,IgE)[12]。 牛初乳中只含有IgG、IgA和IgM[13]。其 中,IgG在 成熟乳中以单体形式存在,而IgA以二聚体或四 聚体形式存在,通过C末端的J链共价链接。分泌 型IgA 二聚体由单体通过分泌组分(SC)和 J链连 接而成[14]。IgM 以 1 000 ku 的五聚体形式存在, 通过二硫键和J链连接。所有免疫球蛋白的基本结构相同,由2个轻链和2个重链通过二硫键连接组成。V代表可变区((Variable heavy chain domain, VH) ,( Variable light chain domain,VL)),C 代表恒 定区((Constant heavy chain domain,CH) ,( Constant light chain domain,CL))。免疫球蛋白的基本结构可以表示为二维的“Y”形 ,见 图 1。IgG分子可以分为 2 个独立的活性域,可变抗原结合域或片段以及恒定 区。抗原结合片段(Fragment antigen-binding,Fab), 通过与抗原的靶标或表位结合的位点,即互补决定 区(Complementarity-Determining,Region,CDR)进行相互作用,见图1(b)。由于 CDR中氨基酸的变异性,IgG 能以高特异性结合各种抗原,因为免疫球蛋白的一般折叠Y形(三级结构)是保守的。可结晶片段(Fragment crystallizable,Fc)是启动免疫反应的效应器,没有抗原结合能力见图1(b)。Fc片段具有某些生物活性,包括被细菌结合蛋白识别、激活补体、通过 上皮细胞传输、被白细胞和上皮细胞上的Fc受体识别等。免疫球蛋白分子有2个翻译后的构象特征,有 助于整体分子稳定性:链内和链间通过半胱氨酸残基形成的共价二硫键以及糖基化,见图1(c)。二硫键是 免疫球蛋白稳定的特征性结构,其能增强蛋白质的稳定性已在多种蛋白体系中得到验证,如黏附分子、结构蛋白等。其他研究将IgG的糖基化与稳定效应系起来,糖基化是Fab域的一个可变特征,但在Fc 域上是一个保守特征[15]。IgG最容易受到酶解的部位是铰链区,它连接了Fab和Fc域。铰链区是IgG 最易弯曲的区域,因为它是IgG唯一的单肽链区域。 抗原结合位点 CH1 CL CH2 CH3 CL 初乳中的免疫球蛋白对保护新生牛犊免受微生物感染至关重要。免疫球蛋白是初乳中的主要抗菌因子, 占初乳总蛋白的70%~80%,而仅占牛乳总蛋白的 1%~2%。
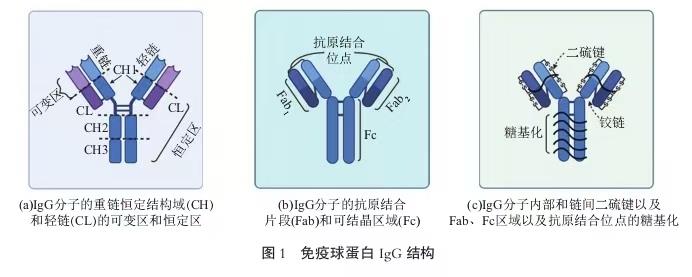
1.2.1 IgG
IgG 抗体能通过其Fc部分与免疫细胞(中性粒 细胞、巨噬细胞等)表面的受体结合,从而增强免疫细胞对病原体的吞噬作用。此外,IgG抗体通过阻断病原体表面的特定表位结合,从而阻止其黏附在宿主细胞或组织上,减少病原体的扩散和感染,起到中和病毒的作用。除了对病原体的抑制作用,IgG抗体 还能直接与细菌的代谢酶结合,从而阻止细菌生长 和繁殖。IgG抗体存在条件下,促使多个细菌聚集成团,不仅有助于免疫细胞的识别和清除,还能进一步减少细菌扩散[16]。综上所述,IgG抗体通过多重机制发挥其免疫调节功能,包括调理素作用、补体固定、防 止病原微生物黏附到内皮内层、通过阻断酶抑制细菌代谢、细菌凝集以及中和毒素和病毒,见图2[17]。IgG 是体内最主要的免疫球蛋白类型,它具有所有免疫球蛋白同种型中最长的血清半衰期。IgG也是目前研究最广泛的免疫球蛋白类别。基于重链恒定区的结构、抗原性和功能差异,特别是CH1和CH3区域 ,已鉴定出4种IgG亚类(IgG1、IgG2、IgG3和IgG4)[17]。 这些IgG亚类的编号是参照生活在西方欧洲健康环境中居住者血液中抗体血清水平的排名顺序(IgG1 > IgG2 > IgG3 > IgG4)来确定的。CH 结构域的差异影响抗体的灵活性和功能性亲和力,其中一些起到与多价抗原的协同作用。抗体Fab和Fc部分的移动性或灵活性主要由CH1结构域和铰链区控制。不同 IgG 亚类表现出不同的功能活性。激活补体级联反应是清除被包被的病原体的重要手段[18]。尽管 IgG4是 唯一不固定补体的亚类,但对(Complement 1q,Ciq) 的亲和力(C1q是补体途径的重要组分,与IgG的 CH2结构域结合)在其他3种IgG亚类成员之间存在差异(IgG3 > IgG1 > IgG2)。在对3类Fcγ受体(Fc gamma receptor,FcγR)( I、II 和 III)的亲和力方面也存在明确差异:IgG1和IgG3与所有3类FcγR结合; IgG4 仅与FcγRII 和FcγRIII 结合,尽管这种结合明显比IgG1的结合弱;IgG2仅与FcγRII结合[19]。
IgG 作为主要的糖蛋白,在新生儿的免疫防御中糖基化 (c)IgG分子内部和链间二硫键以及 Fab、Fc区域以及抗原结合位点的糖基化扮演关键角色。最近研究表明,山羊乳中的IgG的N 糖链结构对于其稳定性和功能至关重要。研究者通过超高效液相色谱四极杆-飞行时间质谱联用技术 (RP-UHPLC Q-Tof MS/MS)对牛乳中的IgG进行了特异性位点N-糖链分析。IgG显示出复杂的糖链结构,主要以双天线形结构存在,并伴随着高程度的岩藻糖化和唾液酸化[20]。这种聚糖特征能够使其作为生物活性成分发挥免疫调节功能。研究发现,牛乳IgG糖基化有助于完善新生儿不断发展的肠道微生物群,从而获得被动免疫保护[21]。
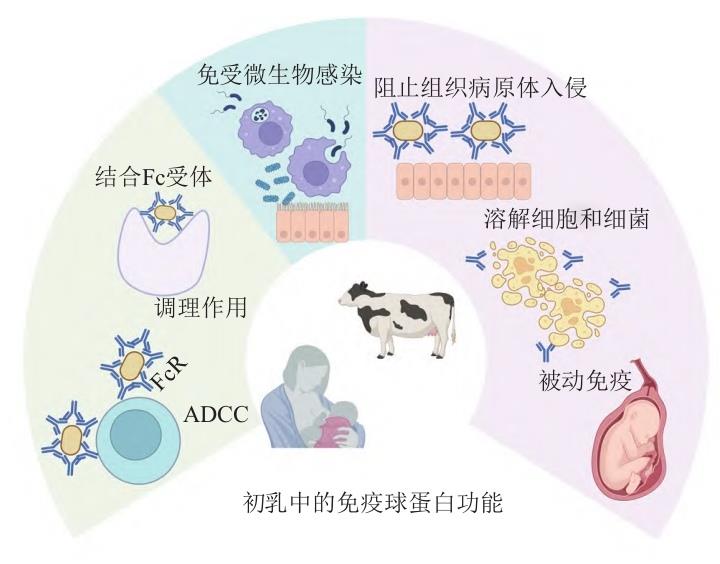
图 2 初乳中免疫球蛋白的功能
IgG 亚类之间也发挥相似的作用,如胎盘转运和参与二次免疫反应。在二次免疫反应中,诱导IgG主要亚类存在偏差。如通常对蛋白质抗原的反应会诱导产生IgG1和IgG3抗体,而与多糖抗原相关的则是 IgG2 和 IgG4 抗体。特定抗原刺激可诱导IgG亚类反应发生偏倚。在疫苗研发时,IgG抗体通过中和毒素、直接清除病原体等机制参与免疫应答。IgG亚类 也通过这种相互作用介导免疫效应。在HIV患者中, IgG3 抗体较IgG1抗体展现出更强的病毒中和效力, 其机制可能涉及抗体铰链区柔性增加、抗体结合表位优化以及病毒包膜寡聚体构象扰动[22]。
1.2.2 IgM
IgM 是B细胞发育过程中首个表达的免疫球蛋白。未成熟的B细胞表面膜结合型IgM(mIgM)与 CD79a 和CD79b 异源二聚体共组装介导抗原识别信号的初级转导,其中Igα(CD79a)和 Igβ(CD79b)胞内段含免疫受体酪氨酸激活基序(Immunoreceptor tyrosine-based activation motif,ITAM)。抗原激活的成熟B细胞分泌以五聚体为主的IgM多聚体,其单 体通过CH4结构域中的二硫键共价连接。五聚体中的多肽链(J链)通过二硫键连接2个单体,介导其向黏膜表面的分泌。通常,单体IgM分子由于其未经历 亲和力成熟而表现出固有的低亲和力,但其五聚体结 构能通过空间拓扑匹配实现多价结合,尤其是对含重复表位抗原可产生亲和力增强效应[23]。 IgM 通过多价结合介导抗原中和并激活补体级联反应发挥免疫作用,其五聚体构型因拓扑匹配显著增强该过程的生物学效应。IgM抗体与初次免疫反应相关,经常用于诊断免疫原或病原体的急性暴露。 IgM 在B细胞初始分化阶段优先表达,其μ重链与未经历体细胞高频突变的VH和VL区域共组装,形成具有广泛反应性的初级抗原识别受体。因此,IgM 抗体因具有广谱表位结合特性,使B淋巴细胞可跨越抗原表位差异性来实现快速活化,这种多反应性机制在初次免疫应答过程中起到了关键作用。低亲和力的IgM抗体也被称为天然抗体,具有双重生物学功能,既能识别病原体表面保守结构(如细菌多 糖)触发补体级联反应,快速清除病原体,又可通过 Fc 受体相互作用参与调节性B细胞的分化,维持免疫耐受平衡。天然抗体会与自身抗原发生反应,但很少导致自身免疫疾病或参与病理过程。致病性自身抗体往往来自经过体细胞突变、高亲和力的IgG 群体[24]。IgM 抗体的产生量小于IgG,但其在补体激活效能和多价中和作用方面具有显著优势,然而这种优势在牛乳中的重要性和贡献机制还存在研究 空白。 研究表明,CD5抗原样蛋白(CD5 antigen-like protein,CD5L)是 IgM 不可或缺的组成部分,以依赖钙离子的方式与J链结合,并通过二硫键与IgM进一 步连接。进一步证实了CD5L不会干扰IgM介导的补体激活,这些结果有助于对IgM更全面的了解,并揭示了J链在免疫系统中的作用[25]。在五聚体或六聚体IgM中,Fc区域在溶液中采用平面构象,直到抗原结合引发构象变化并暴露补体结合位点。IgE-Fc 在溶液中主要采用急剧弯曲的构象,但不同受体的结合受弯曲程度控制,并且受体结合位点之间存在变构通讯[26]。此 外,IgM具有独特铰链区,在南极鱼类中发现,IgM具有一个特别长的铰链区,影响IgM功能,包括其与抗原的结合能力[27]。IgM不仅是宿主抵御感染的第一道防线,还在免疫调节和免疫耐受中起着重要作用。多年来,人们初步认为IgM调节免疫反应是通过与抗原结合和激活补体系统来发挥作用。 但随着对IgM的Fc受体的发现,目前普遍认为IgM 也可通过其受体发挥作用,并在B细胞发育和活化、 体液免疫反应、宿主防御和免疫耐受中扮演重要角色[28]。综上所述,IgM在免疫应答中的作用具有多样性和复杂性,贯穿从早期细菌感染、补体系统的激活, 再到黏膜免疫和天然免疫全过程。
1.2.3 IgA
相比之下,分泌型IgA(Secretory immunoglobulin A,sIgA)虽然无法直接激活补体途径或调理素细菌, 但能够凝集抗原,中和病毒和细菌毒素,并防止肠道致病菌黏附到黏膜上皮细胞上。IgA主要参与局部免疫,sIgA在保护免受GI感染方面起主要作用。IgA 对于保护黏膜表面免受毒素、病毒和细菌的侵害至关重要,通过双重机制维护黏膜免疫稳态:其抗原结片段介导特异性病原体中和以及J链通过与聚合物共价连接产生空间位阻效应阻断微生物黏附分子与上皮受体相互作用,这种双重防护体系对于消化道免疫防御具有不可替代的作用,见图2[29]。此外,细胞内IgA在预防细菌或病毒感染、病理过程中也发挥重要作用[30]。sIgA的多聚体结构具有重要的功能优势, 例如,多聚体形式IgA(Polymeric immunoglobulin A, pIgA)对艰难梭菌毒素A诱导的肠上皮细胞损伤方面展现出比单体IgA更有效的中和能力。虽然IgA在黏膜表面的补体固定不是主要的效应机制,但IgA受体与中性粒细胞表面表达的Fcα受体(FcαRI,CD89) 结合,通过脱颗粒和活性氧释放,在黏膜感染局部介导抗体依赖性细胞介导的细胞毒性(Antibody dependent cellular cytotoxicity,ADCC),从而靶向清除被IgA包被的病原体。如上所述,IgA的糖基化修饰在病原体清除中发挥关键作用,通过糖-凝集素相互作用特异性捕获包膜病原体,形成免疫复合物以促进其清除。sIgA被证实可也能通过树突状细胞表面的受体介导抗原内吞,这种跨膜转运机制使肠道固有层中的树突状细胞将sIgA-抗原复合物提呈至T细 胞,诱发记忆B细胞形成,放大黏膜免疫。这些特性使得IgA成为黏膜免疫防御的关键组成部分,在防止病原体穿透黏膜屏障和调节局部免疫反应中发挥着重要作用,见图2[31]。
人类中存在IgA的2种亚型,即IgA1和 IgA2, 其中IgA1亚型占主导地位。IgA1与IgA2的不同之处在于铰链区额外有13个氨基酸,由2个重复的氨基酸序列组成。这些序列具有丝氨酸和苏氨酸的多个O-糖基化位点。IgA2在铰链区缺乏O-糖基化位点, 仅显示由葡萄糖胺(GlcN)修饰的N-糖基化位点, 并且表现出较低水平的唾液酸化。值得注意的是, IgA1 的糖基化具有高度异质性[32]。与 IgG类似,IgA 的糖链结构(如高甘露糖型或复合型)影响与Fcα受体(FcαRI)的结合能力,进而调节炎症反应和黏膜免疫。为了维持黏膜免疫的微妙平衡,分泌型IgA具有重要的效应和调节功能:(1)肠管腔侧的免疫排斥, 阻断病原微生物的定植和渗透,降低黏膜屏障的慢性炎症 ;(2)通过诱导抗原特异性效应反应(包括吞噬 作用、抗体依赖性细胞毒性)中和病原体,实现超氧化物的产生、炎症介质和细胞因子的释放以及抗原呈递。最近研究表明,IgA抑制过敏反应的抗炎功能是通过对肥大细胞活化抑制来实现的,并且取决于IgA 的唾液酸修饰[32]。IgA在黏膜免疫、抗感染和肿瘤治疗中展现出独特优势。未来研究需进一步解析Fc受体互作机制,并探索基于IgA结构的精准免疫调控策略。
2 初乳中免疫球蛋白的应用潜力
初乳被确定为新型食品和药物配方中具有前景的成分,应用于包括口腔卫生、胃肠功能、生长因子 (食品补充剂和非处方药品)、婴儿配方奶粉、特定药 物、自身免疫疾病、类风湿性关节炎、多发性硬化症和化妆品[33-34]中。当前市场上的商业化免疫球蛋白产品是通过收集初乳、脱脂、在保持生物活性的条件下 通过巴氏杀菌和干燥制成的。产品形式包括喷雾干燥和冻干粉末,以及过滤的初乳清液或浓缩物。干燥 产品包括全脂乳粉、脱脂粉、脱脂初乳蛋白浓缩物和 初乳清浓缩物。一些潜在的促进健康的免疫球蛋白成分正处于专有开发或临床研究中[35]。来自未免疫 奶牛的牛初乳在人类健康中的预防或治疗用途并不罕见,可以追溯到几十年前。事实上,饮用牛乳能够 预防感染在人类历史上已得到多个世纪的验证。然 而最近,通过现代膜分离和加工技术可实现的活性 IgG 的大规模富集或分离刺激了营养保健食品和膳食补充剂市场的快速增长[36]。因此,尽管存在分类学问题,但与人源制剂的潜在用途相比,用于人类健康的反刍动物衍生抗体的开发占主导地位。牛乳中的免疫球蛋白已被用于婴幼儿配方奶粉和其他食品的免疫补充剂。在竞争激烈且有价值的含IgG产品的国际市场上,产品通常根据IgG含量定价,因此这些产品声称的IgG含量精确性非常重要。
研究表明,富含免疫球蛋白的免疫制品可有效增强对抗轮状病毒(腹泻)、大肠杆菌(腹泻)、白色念珠菌(念珠菌病)、艰难梭菌(结肠炎、发热)、志贺 氏菌(痢疾)、变形链球菌(龋齿)、隐孢子虫(腹泻) 和幽门螺杆菌(消化性溃疡)等的感染,预防它们引起的人类和动物疾病[39]。基于上述作用机制,牛初乳和乳清产品(宣称增强免疫系统功能)通常与商品化分离IgG产品具有等同性,其健康声明的科学依据主 要为可改善GI及口腔健康。群体健康认知提高使得自身免疫状态得到关注成为国际公共卫生领域的焦点议题,主要反映在增强免疫力产品的销售提升,其中牛乳为许多产品提供了基础,尤其是针对于早产儿、婴儿、 幼儿和一些学龄儿童[40]。这种趋势建立在人们相信口服IgG产品是安全的,并且为消费者提供抗GI感染的措施保护。消费者对这种IgG增值乳清蛋白产品的接受始于亚洲,现在已经传播到欧洲和美国,目标是体育营养、婴儿配方奶粉、膳食补充剂和生理功能食品。 尽管牛初乳含有针对特定肠道病原体的IgG,但初乳的产量和质量受到接触病原体、代谢健康、产前 管理、胎次等因素的影响,不能提供优质的初乳从而直接影响IgG含量[41]。这些无法控制的影响因素导致牛初乳无法为人类提供最佳保护 。克服正常初乳存在的这种限制的策略之一是登记奶牛的产犊时间、 健康状况、采样时间等信息。根据正确的采样方式, 将初乳收集限制在分娩的6 h内(此时IgG含量非常 高),或者通过提高分离技术来制备IgG。提高正常牛初乳临床效力的进一步改进措施是生产和提供超免疫乳[42]。来自用人类肠致病微生物免疫的牛IgG强化乳已被证明具有显著保护作用,其作用机制主要通过调节微生物对肠道上皮细胞的黏附来实现。近期,针对免疫活性乳制品(含特异性IgG浓缩物)在人体感染性疾病治疗中的应用相关综述表明,基于体外和动物实验研究显示,该类干预措施在细菌性GI感染、口腔病原体定植、寄生虫侵染及急性病毒性感染等短期治疗场景中展现出暂时性疗效。
3 免疫球蛋白的消化和吸收特性
腹泻是发展中国家5岁以下儿童疾病和死亡的主要原因。提供针对肠道病原体的口服特异性抗体补充剂是降低疾病发生的一种潜在治疗措施。为预防感染,补充免疫球蛋白理想上需要抵抗儿童GI降解。因此,了解免疫球蛋白在GI中的消化吸收特性对于理解被动免疫的机制和效果至关重要。GI中免疫球蛋白的消化吸收不仅影响机体的免疫保护作用, 还会对营养吸收产生积极影响。此外,定植在肠道中的免疫球蛋白通过结合微生物来发挥健康益处,从而 防止肠道细菌移位和继发性炎症[44]。研究表明,特别是在初乳中的免疫球蛋白,在保护GI中扮演重要的生物学角色,主要包括保护婴儿免受肠道感染[45]、增强肠道屏障完整性[46]和调节肠道免疫反应等方面[47]。 IgA 分子的分泌片段组分使其具有更显著的抗蛋白酶消化能力[48]。摄取后,乳源IgG遇到被吞入的呼吸道病原体和吸入的过敏原时可部分阻止这些病原体和过敏原进入身体,从而减少感染和过敏反应,特别是在乳汁被重吸收并进入鼻咽部和咽淋巴环的扁桃体时,能在这些部位进行免疫调节,包括调节性T细胞 (Treg)和免疫球蛋白A(IgA)的产生,见图3(a)。在小肠中,IgG也可通过阻止病原体黏附到上皮表面来排除病原体,但也通过Fc受体促进免疫复合物的摄取,包括与病原体结合的IgA,从而引发免疫反应和诱导局部IgA的产生,见图3(b)。众所周知,在人体 GI 中,牛乳免疫球蛋白被蛋白酶,如胃蛋白酶、胰蛋 白酶、糜蛋白酶、羧肽酶和弹性蛋白酶分解成F(ab')2、 Fab 和 Fc 片段。IgG消化为Fab二聚体(约100 ku) 和单体Fab片段(约 50 ku),但这种酶消化的敏感性因IgG亚型和物种而异。如牛乳中的IgG1比IgG2 更容易被胃蛋白酶水解。牛IgG与兔或人类IgG相 比,在蛋白酶消化中更稳定。即使在IgG经过蛋白酶消化后,只要Fab二聚体和单体片段没有变性,它们 就仍然保持结合和中和抗原的活性。体外试验对牛源性IgG1 的蛋白酶特性研究表明,Fab域完整且具有反应活性。此外,调节酶消化过程中的反应时间、pH值和温度将产生各种不同的活性域片段 [49]。然而,所产生的F(ab')2 和Fab 片段在口腔和食道中仍保持大部分天然活性,完整的免疫球蛋白、F(ab')2和其他抗体正在被用于靶向治疗制剂的开发。
在小肠中,免疫球蛋白会进一步被胰酶消化。其 中,胰蛋白酶优先降解牛IgG1重链的铰链区而不是 IgM,而另一种酶糜蛋白酶则更倾向于水解IgM的 Fc μ 结构域而不是IgG,IgG2则更容易被胰蛋白酶水解[49]。与牛乳中其他乳清蛋白相比,免疫球蛋白更能抵抗GI消化。在肠道中,其他主要乳清蛋白的消化特性是α-乳白蛋白被迅速消化,而β-乳球蛋白 消化较慢[50]。IgG 在GI消化模型中显示出蛋白酶抗性,这种特性使其成为GI转运过程中半衰期中最长的成分,与其他主要乳清蛋白相比,IgG为新生儿提供的氨基酸比例最小[51]。在体外将IgA和IgG与 小羊的小肠内容物共同孵育,结果显示IgA比IgG具 有更强的抗消化性。在成年人摄入牛乳清蛋白浓缩 物时,通过径向免疫扩散从空肠流出物中检测到大约59%的IgG和IgM,而在回肠中检测到19%。这些 免疫球蛋白消化的估计值与成年人摄入的牛乳清蛋 白浓缩物消化的估计值相比较,在空肠末端大约完 成42%,在回肠末端完成93%,再次强调了免疫球蛋 白在GI中较强的抗消化能力,见图3[52]。此外,在结肠中,IgG可防止脂多糖(LPS)的泄露,可改变微物群落的组成和产生短链脂肪酸(SCFA),并且可防止病原体黏附,见图3(c)。许多早期发表的关于口服IgG消化性的临床研究均是在婴儿群体中进行的。 HILPERT 等[53]研究表明,在接受针对肠致病性大肠杆菌(Eneropathogenic Escherichia coli,EPEC)免疫的牛初乳来源免疫球蛋白的婴儿粪便中,存在EPEC 的特异性IgG或其活性片段。与此一致的是,在接 受了口服初乳免疫球蛋白的轮状病毒儿童的粪便 中,可检测到抗轮状病毒免疫球蛋白活性[54]。这些数据表明,IgG能够通过GI的传递并保持结构完整性。此外,在另一项研究中,6名健康的、未成熟的、 配方奶喂养的婴儿在连续5 d内每天分次摄入10% 的人类免疫血清球蛋白(主要为IgG),剂量为每天 1~8 mL/kg。在 24 h 内,粪便中IgG的存活率在摄入原始IgG的4%~12%之间。虽然受试者粪便中活性 IgG 的存活情况有差异,但增加的口服剂量与每天排出更多的IgG相关。这些试验结果表明,婴儿口 服以IgG为主要原料的婴幼儿配方奶粉并没有出现慢性腹泻、体质量减少和吸收不良等反应[49]。经双盲随机对照试验验证,通过口服的牛源IgG通过GI 上皮细胞FcRn受体介导的胞吞转运,能够在肠黏膜层形成免疫保护,防止致病性大肠杆菌黏附和穿透,减少新生儿感染的风险,见图3。在一项更大规模的随机对照临床试验中,给予不能母乳喂养的低出生体质量婴儿每天600mg的血清来源的人类IgA (73%)和 IgG(26%)。测试组喂养含有IgA和IgG的混合物(模拟母乳中的IgA和IgG的比例)(n=91), 对照组(n=88)喂养相同的配方奶粉或母乳制剂,但不包含血清来源的IgA和IgG。与对照组(6例)相比,接受口服IgA-IgG的婴儿患坏死性小肠结肠炎的病例较少(0例),并且在粪便中检测到大量的完 整IgA和IgG[55]。
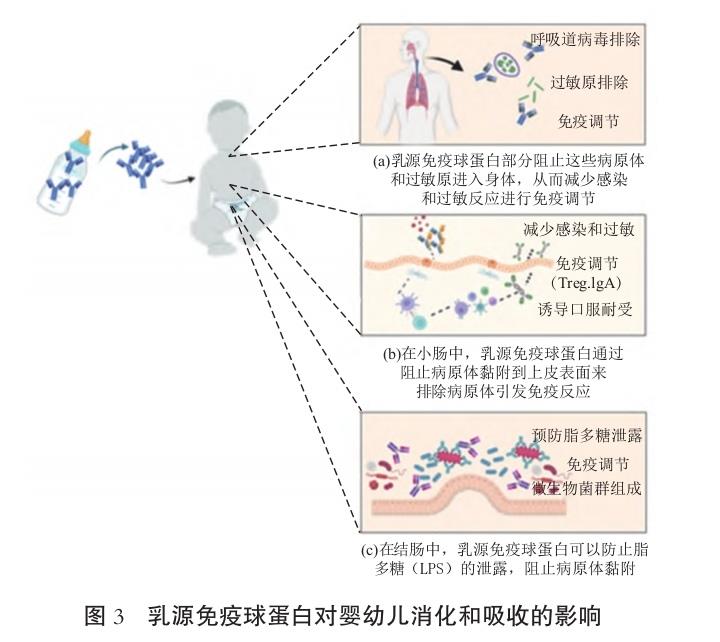
3.1 免疫球蛋白稳定性
蛋白质的生物活性表达高度依赖于其三维折叠结构,天然状态被视为时间是平均的、热力学上最稳定的构象。通常,蛋白质变性可分为部分变性和广泛变性。部分变性虽然导致其天然构象受到干扰,但仍保留一定功能,而广泛变性则会最终导致蛋白质完全丧失生理功能。因此,蛋白质变性导致多肽链的展 开,球状结构伸展开使得-SH键暴露,最终导致不可 逆的高阶聚集和沉淀[59]。生物活性蛋白质的构象完整性对变性的抗性随着单一蛋白质分离物在食品治疗领域的趋势日益增强,这一特性对IgG富集食品和 补充剂的生理功效尤为重要。无论牛IgG的来源如何(完整的初乳、超免疫乳、免疫球蛋白分离物等), 在生产、储存和GI转运等环节维持其Fab和Fc结构域的空间构象完整性,是确保其抗原结合活性与免疫调节功能的关键因素。初乳免疫球蛋白相对热敏感,为了保持其构象完整性和生物活性,在乳制品加工过程中,必须尽量减少牛乳蛋白暴露于热环境中。 值得注意的是,在热变性条件下,通过免疫分析法 (Enzyme-linked immunosorbent assecy,ELISA)检测的抗体免疫反应性可能不一定等同于特定病原体中和试验定量的抗原结合活性,这一现象表明,这2种特性会以不同的方式受到热变性条件的影响[11]。不同的温度和湿度均是影响IgG的重要因素,导致IgG活性降低。研究发现,在低温条件下对IgG免疫活性物质未造成显著影响,而在高温条件下(65 ℃)其活性损失率高达77.2%。因此,冷冻干燥加工过程虽然对活性免疫球蛋白造成一定程度损失,但与喷雾干燥处理相比(0.0143 mg/mL),冷冻干燥能更有效地保留生物活性物质,含量约0.2288 mg/mL[60]。
近期的研究聚焦于牛IgG在模拟体系(磷酸盐 缓冲液)及天然乳基质(牛乳)中的构象稳定性方面,以评估其在长期冷冻储存、分离和加工(热、pH 和压力)各种环境应激条件下的活性保留率和结构 完整性[61]。现有的研究采用多原理分析技术评估了 IgG 的稳定性,由于检测方法的异质性导致跨研究数 据呈现离散性。然而,通常表明,IgG的结构和功能 在大约65 ℃以上的热处理中是不稳定的,因此传统 的巴氏杀菌条件对保护IgG结构完整性有一定的局限性。免疫检测分析揭示了IgG内部可能存在轻微的结构稳定性差异。不同物种来源IgG的Fab/Fc结 构域在加热、pH和蛋白酶条件下,虽呈现构象柔性, 但仍然保留其核心结构[62]。因此,目前的研究结果表明 ,牛 IgG制剂的免疫预防或治疗作用在人类口服给药途径下具有良好的生物相容性。
3.2 影响免疫球蛋白消化吸收的因素
口服免疫球蛋白制品(人血清、牛初乳和血清 来源免疫球蛋白)对各种肠道微生物(如大肠杆菌 和轮状病毒等)引发的感染性腹泻展现出显著的治疗效果。在婴儿、儿童和成人中显示,口服IgG后的粪便回收率存在显著的年龄依赖性差异:婴儿、儿童 (0.5%~10%)<成 人(25%)。通常认为,只有当 Fab 结构完整且未被酸性 pH或消化蛋白水解酶完全变性时,IgG 才能通过构象表位识别作用与GI内的抗原结合[63]。来源于人血清以及牛初乳和血清的免疫球 蛋白制剂在模拟GI环境中维持结构完整性,肠道各段均显示出较长时间的驻留。免疫球蛋白制剂中, IgG 是主要成分,而 IgA和IgM的含量通常较低。 蛋白质熔化温度(Melting temperature,TM)是指蛋白质展开或变性导致其丧失其三维结构的特征温 度。TM是生化测量指标,用于在不同情况下探测蛋 白质结构。IgG的热稳定性与pH值相关:较低的pH 值(2.8~3.4)对应较低的TM值(43.7~53.6 ℃ ),较 高 的pH值(5.0~7.5)对应较高的TM值(67.5~68 ℃)。 即使在2.8~3.4 的酸性pH范围内,TM值也不会低于体温37 ℃,这就解释了免疫球蛋白在消化道中能够维持结构完整性[64]的原因。在缓冲液中添加糖 类,如海藻糖和蔗糖,可提高所有IgG的TM值 ,并 将pH值维持在4.0~8.0之间,这表明GI内其他环境因素,如饮食分子,可能会增加免疫球蛋白的稳定性[65],但盐分含量并不影响IgG稳定性。IgG的CH2结构域,在暴露于pH 2.0后呈现出热稳定性的异常增加,其机制源于pH诱导IgG的β折叠网络重构和盐桥重组来实现结构稳定性。利用pH梯度洗脱纯化的IgG分子仍具有完整的抗原结合活 性,其免疫反应性未发生显著改变。实际上,某些静脉注射免疫球蛋白制剂,通过低pH纯化工艺处理, 能增强体外和体内结合抗原的能力。这些研究强调 了IgG在生理温度协同酸性微环境中仍可保持其构象稳定性。胃中酸性环境通过诱导IgG分子构象重排,能增加其在GI中的生物稳定性和抗蛋白酶水解能力。
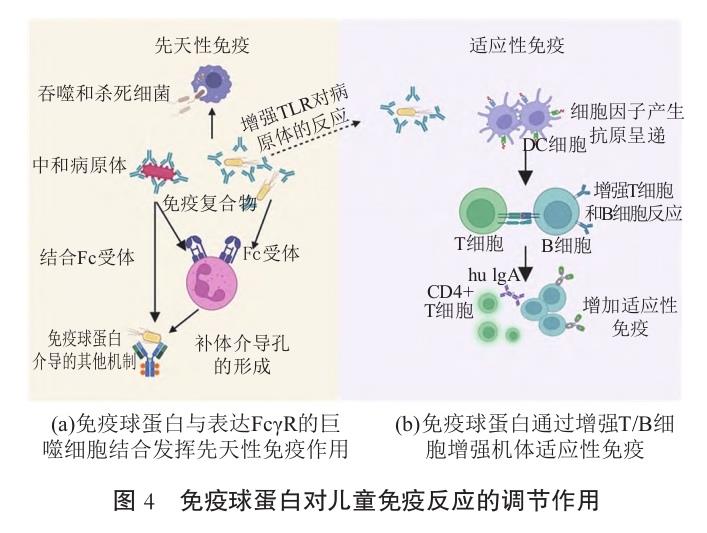
4 免疫球蛋白对儿童免疫发育的影响
免疫球蛋白可通过与病原体表面的抗原结合起到中和保护作用。当病原体侵入后,免疫球蛋白可迅速调动机体内其他免疫细胞,如巨噬细胞、中性粒细 胞、树突状细胞等,进一步起到清除作用。同时发现儿童的炎症系统相比成人的较为剧烈,容易引发其他器官发生疾病,导致免疫系统紊乱和炎症系统失控。 免疫球蛋白还能激活补体系统,进一步来消灭病原 体,见图4。 免疫球蛋白对免疫发育的影响作用主要通过以下机制来发挥作用。(1)免疫球蛋白抗体与自身免疫系统中的其他抗体协同发挥作用,增强对病原微生物的识别和清除能力;(2)免疫球蛋白的抗体直接中和病原微生物表面毒素,阻止其进入机体破坏机体细 胞。在与微生物抗原结合后,减弱病原微生物活性, 使其更容易被免疫系统激活以及进一步清除;(3)摄入免疫球蛋白能阻断体内炎症信号转导,抑制炎症细胞的活化和迁移,调控T淋巴细胞亚群水平,从而改善机体免疫功能,见图4。免疫球蛋白在儿童免疫发育中具有独特且多方面的作用,尤其体现在被动免疫、免疫调节和病理机制干预方面。免疫球蛋白在参与抵抗儿童呼吸道和消化道感染方面发挥重要作用,尤其是为婴儿期和学龄前儿童提供被动免疫方面。研究发现,免疫球蛋白作为治疗儿童川崎病的辅助手段,其治疗的优势主要是改善调理素作用和吞噬作用[66]。另一项研究表明,免疫球蛋白干预对面部异常综合征儿童在治疗早期具有缓解作用,呼吸道细菌感染频率有所下降[67]。免疫球蛋白治疗已被证实对儿童自身免疫疾病(免疫性血小板减少症)安全有效,通常表现出耐受性良好。其治疗主要通过中和自身抗体,调节巨噬细胞Fcγ受体的表达,抑制炎症性通路来发挥免疫调节作用[68]。此外,发现儿童通过疫苗接种产生的病原体特异性的IgG(如抗流感病毒 H3N2中和抗体)可提供直接保护,通过表位交叉反应抑制病原体感染,从而增强对病原体的适应性免疫应答[69]。因此,免疫球蛋白在儿童免疫发育中不仅是效应分子,更是免疫稳态的动态调节媒介,其作用涉及体液免疫、自身免疫反应和过敏反应等多个环节。 其动态变化与儿童特有的免疫缺陷、自身免疫疾病密切相关,可为早期诊断和靶向治疗提供依据。
5 总结和展望
牛乳是免疫球蛋白的丰富来源。这些分泌物是通过进化发展而来的,以确保被动免疫从母体到后代 的同源传递。从母体传给后代的免疫球蛋白,无论是通过经胎盘转移还是通过初乳的摄入,均可在母体抗原暴露史和新生儿的被动免疫保护之间形成重要联 系。这种免疫学联系不仅包括免疫球蛋白的垂直传 递,其乳腺分泌物中也已验证存在多种协同作用的免 疫调节因子。牛乳中免疫球蛋白的生物活性特征,为开发跨物种被动免疫制剂(如口服免疫调节剂)提供了分子基础。研究表明,牛初乳中的非特异性免疫组分通过跨物种FcRn保守结构域介导,在无需抗原预暴露的条件下即可实现异源被动免疫转移。基于免疫球蛋白构象稳定性优化与肠道靶向递送系统制剂技术的创新,通过疫苗佐剂效应强化、热稳定性增强 及加工敏感性降低,能显著提升牛乳生物制品的临床转化潜力。虽然免疫球蛋白从母体被动免疫传递机 制及其在新生儿中的作用已得到充分阐述,但乳源免疫组分在儿童和成年动物模型中的生物活性机制,特别是FcRn非依赖性转运路径及肠道菌群互作效应仍亟待深入解析。对宣称的添加IgG的婴幼儿配方奶粉能够改善儿童免疫功能、减轻炎症反应等方面的研究日益受到临床重视,深入了解免疫球蛋白对儿童免疫发育的影响将有利于富含IgG婴幼儿配方奶粉的开发和利用,为儿童免疫发育以及免疫状态的预判和预防提供健康指导。
参 考 文 献:
[1] SUDARSHAN M,DIVYA S V,JYOTHIBAI P,et al. N-Glycoprofiling of immunoglobulin G and lactoperoxidase from sheep milk using LC MS/MS[J]. International Dairy Journal,2023,140: 105582.
[2] LOPEZ A,HEINRICHS A. Invited review: The importance of colostrum in the newborn dairy calf[J].Journal of Dairy Science,2022,105(4): 2733-2749.
[3] PROSKURINA L,ENNS E,SIMAKOVA M,et al. Cell-mediated transfer of immune protection factors to newborn calves[J].E3S Web of Conferences,2020,203(1): 01030.
[4] WESTRŐM B,ARÉVALO S E,PIERZYNOWSKA K,et al. The immature gut barrier and its importance in establishing immunity in newborn mammals[J]. Frontiers in Immunology,2020,11: 1153.
[5] LOZANO N A,LOZANO A,MARINI V,et al. Expression of FcRn receptor in placental tissue and its relationship with IgG levels in term and preterm newborns[J]. American Journal of Reproductive Immunology, 2018,80(3): 12972.
[6] YOSHIDA M,MASUDA A,KUO T T,et al. IgG transport across mucosal barriers by neonatal Fc receptor for IgG and mucosal immunity[J]. Springer Seminars in Immunopathology,2006,28(4): 397-403.
[7] BAKER K,QIAO S W,KUO T,et al. Immune and non-immune functions of the(not so)neonatal Fc receptor,FcRn[J]. Seminars in Immunopathology,2009,31(2): 223-236.
[8] QIAN S,ZHANG D,YANG Z,et al. The role of immunoglobulin transport receptor,neonatal Fc receptor in mucosal infection and immunity and therapeutic intervention[J]. International Immunopharmacology, 2024,138: 112583.
[9] ARSLAN A,DUMAN H,KAPLAN M,et al. Determining total protein and bioactive protein concentrations in bovine colostrum[J].Journal of Visualized Experiments(JoVE),2021,178: 63001.
[10] JAWHARA S. Can drinking microfiltered raw immune milk from cows immunized against SARS-CoV-2 provide short-term protection against COVID-19[J]. Frontiers in Immunology,2020,11:1888.
[11] GAPPER L W,COPESTAKE D E J,OTTER D E,et al. Analysis of bovine immunoglobulin G in milk,colostrum and dietary supplements: a review[J]. Analytical and Bioanalytical Chemistry,2007,389(1): 93 109.
[12] MARIE A M,ABIDALKAREEM I H. Study of the Levels of immunoglobulin G and complement 3 in gestational diabetes and their newborn[J]. International Journal of Medical Science and Clinical Research Studies,2023,3(9): 2066-2074.
[13] SCHROEDER H W,CAVACINI L. Structure and function of immunoglobulins[J]. Journal of Allergy and Clinical Immunology, 2010,125(2): S41-S52.
[14] JOHANSEN,BRAATHEN,BRANDTZAEG. Role of J Chain in secretory immunoglobulin formation[J]. Scandinavian Journal of Immunology,2010,52(3): 240-248. [15] ARNOLD M J J,CARL G K,KIMBERLY W,et al. Contributions of a disulfide bond to the structure,stability,and dimerization of human IgG1 antibody CH3 domain[J]. Protein Science,2008,17(1): 95-106.
[16] NISHIYAMA A,NOGIMORI T,MASUTA Y,et al. Cross-reactive Fc-Mediated antibody responses to influenza HA stem region in human sera following seasonal vaccination[J]. Vaccines,2025,13(2): 140.
[17] BOURNAZOS S,GUPTA A,RAVETCH J V. The role of IgG Fc receptors in antibody-dependent enhancement[J]. Nature Reviews Immunology,2020,20(10): 633-643.[18] CAVACINI L A,KUHRT D,DUVAL M,et al. Binding and neutralization activity of human IgG1 and IgG3 from serum of HIV infected individuals[J]. Aids research and human retroviruses,2003,19 (9): 785-792.
[19] PARLOWSKY T,WELZEL J,AMAGAI M,et al. Neonatal pemphigus vulgaris: IgG4 autoantibodies to desmoglein 3 induce skin blisters in newborns[J]. Journal of the American Academy of Dermatology,2003, 48(4): 623-625.
[20] GNANESH K B S,LIJINA P,JINESH P,et al. N-Glycoprofiling of immunoglobulin G and lactoferrin with site-specificity from goat milk using RP-UHPLC MS/MS[J]. Food Chemistry,2022,383: 132376.
[21] FEENEY S,GERLACH J Q,SLATTERY H,et al. Lectin microarray profiling and monosaccharide analysis of bovine milk immunoglobulin G oligosaccharides during the first 10 days of lactation[J]. Food Science & Nutrition,2019,7(5): 1564-1572.
[22] RICHARDSON S I,AYRES F,MANAMELA N P,et al. HIV broadly neutralizing antibodies expressed as IgG3 preserve neutralization potency and show improved Fc effector function[J]. Frontiers in Immunology, 2021,12: 733958.
[23] LI Y,WANG G,LI N,et al. Structural insights into immunoglobulin M[J]. Science,2020,367(6481): 1014-1017.
[24] BOES M. Role of natural and immune IgM antibodies in immune responses[J]. Molecular Immunology,2000,37(18): 1141-1149.
[25] WANG Y,SU C,JI C,et al. CD5L associates with IgM via the J chain[J]. Nature Communications,2024,15(1): 8397. Vol.53,No.11 2025(total 420) 29 乳功能成分营养与健康专栏 Column
[26] CALVERT R A,NYAMBOYA R A,BEAVIL A J,et al. The evolution of flexibility and function in the Fc domains of IgM,IgY,and IgE[J]. Frontiers in Immunology,2024,15: 1389494.
[27] AMETRANO A,MIRANDA B,MORETTA R,et al. A structural peculiarity of antarctic fish IgM drives the generation of an engineered mAb by CRISPR/Cas9[J]. Frontiers in Bioengineering and Biotechnology,2024,12: 1315633.
[28] LIU J,WANG Y,XIONG E,et al. Role of the IgM Fc receptor in immunity and tolerance[J]. Frontiers in Immunology,2019,10: 529.
[29] KUMAR N,ARTHUR C P,CIFERRI C,et al. Structure of the secretory immunoglobulin a core[J]. Science,2020,367(6481): 1008 1014.
[30] WOOF J M,MESTECKY J. Mucosal immunoglobulins[J]. Immunological Reviews,2005,206(1): 64.
[31] VILLOT C,CHEN Y,PEDGERACHNY K,et al. Early supplementation of Saccharomyces cerevisiae boulardii CNCM I-1079 in newborn dairy calves increases IgA production in the intestine at 1 week of age[J]. Journal of Dairy Science,2020,103(9): 8615-8628.
[32] SCHEURER S,JUNKER A C,HE C,et al. The role of IgA in the manifestation and prevention of allergic immune responses[J]. Current Allergy and Asthma Reports,2023,23(10): 589-600.
[33] PLAYFORD R,WEISER M. Bovine colostrum: Its constituents and uses[J]. Nutrients,2021,13(1): 265.
[34] HAN G,KIM H,KIM D E,et al. The potential of bovine colostrum derived exosomes to repair aged and damaged skin cells[J]. Pharmaceutics, 2022,14(2): 307. [35] KAPLAN M,ARSLAN A,DUMAN H,et al. Production of bovine colostrum for human consumption to improve health[J]. Frontiers in Pharmacology,2021,12: 796824. [36] FENG L,SONG Y,BASURAY S,et al. IgG-BSA separation and purification by internally staged ultrafiltration[J]. Separation and Purification Technology,2025,354: 129245.
[37] 刘影,郝岩. 牛初乳类产品的安全性毒理学评价[J]. 中国乳业,2018, (10): 46-51.
[38] 金洪伟,陈映新. 高含量IgG牛初乳胶囊的安全性研究[J]. 中国乳业, 2018,( 11): 59-64.
[39] AWAD H A,IMAM S S,ABOUSHADY N M,et al. Gut priming with oral bovine colostrum for preterm neonates: a randomized control trial[J]. QJM: An International Journal of Medicine,2020,113 (Supplement_1): 063-025.
[40] TZIPORI S,ROBERTON D,CHAPMAN C. Remission of diarrhoea due to cryptosporidiosis in an immunodeficient child treated with hyperimmune bovine colostrum[J]. British Medical Journal,1986,293 (6557): 1276-1277.
[41] GOI A,COSTA A,PENASA M,et al. Genetic associations of colostrum yield with antibody concentrations and gross composition for selective breeding in Italian Holsteins[J]. Journal of Dairy Science,2025,108(3): 2697-2709.
[42] VIDARSSON G,DEKKERS G,RISPENS T. IgG Subclasses and allotypes: from structure to effector functions[J]. Frontiers in Immunology,2014,5: 520.
[43] KANAGARATHAM C,EL ANSARI Y S,LEWIS O L,et al. IgE and IgG antibodies as regulators of mast cell and basophil functions in food allergy[J]. Frontiers in Immunology,2020,11: 603050. 30
[44] VAN DEN ABBEELE P,DETZEL C,ROSE A,et al. Serum-derived bovine immunoglobulin stimulates SCFA production by specific microbes in the ex vivo SIFR(®)technology[J]. Microorganisms,2023,11(3): 659.
[45] MAOZHEN Z,XI L,MENG Y,et al. Probiotics induce intestinal IgA secretion in weanling mice potentially through promoting intestinal APRIL expression and modulating the gut microbiota composition[J]. Food & Function,2024,15(9): 4862-4873.
[46] KIM S J,LEE H K,KANG K S,et al. Korean red ginseng polysaccharides enhance intestinal IgA production and barrier function via peyer’s patch activation in mice[J]. Nutrients,2024,16(22): 3816.
[47] SINISCALCO E R,WILLIAMS A,EISENBARTH S C. All roads lead to IgA: mapping the many pathways of IgA induction in the gut[J]. Immunological Reviews,2024,326(1): 66-82.
[48] LUEANGSAKULTHAI J,SAH B N P,SCOTTOLINE B P,et al. Survival of recombinant monoclonal and naturally-occurring human milk immunoglobulins A and G specific to respiratory syncytial virus F protein across simulated human infant gastrointestinal digestion[J]. Journal of Functional Foods,2020,73: 104115.
[49] JASION V S,BURNETT B P. Survival and digestibility of orally administered immunoglobulin preparations containing IgG through the gastrointestinal tract in humans[J]. Nutrition Journal,2015,14: 22.
[50] LOVEDAY S M. Protein digestion and absorption: the influence of food processing[J]. Nutrition Research Reviews,2023,36(2): 544-559.
[51] SAADI S,MAKHLOUF C,NACER N E,et al. Whey proteins as multifunctional food materials: recent advancements in hydrolysis, separation,and peptidomimetic approaches[J].Comprehensive Reviews in Food Science and Food Safety,2024,23(1): 13288.
[52] HURLEY W L,THEIL P K. Perspectives on immunoglobulins in colostrum and milk[J]. Nutrients,2011,3(4): 442-474.
[53] HILPERT H,GERBER H,PEYER E D,et al. Gastrointestinal passage of bovine anti-E. coli milk immunoglobulins (Ig) in infants[J]. Nestle Research News,1976,75: 134-138.
[54] BRISTOL L S,DUHAMEL G E,ZINCKGRAF J W,et al. Effect of passive antibodies derived from rotavirus-like particles on neonatal calf diarrhea caused by rotavirus in an oral challenge model[J]. Journal of Dairy Science,2021,104(11): 11922-11930.
[55] EIBL M M,WOLF H M,FüRNKRANZ H,et al. Prevention of necrotizing enterocolitis in low-birth-weight infants by IgA-IgG feeding[J]. New England Journal of Medicine,1988,319(1): 1-7.
[56] ROOS N,S M,BENAMOUZIG R,et al. 15N-labeled immunoglobulins from bovine colostrum are partially resistant to digestion in human intestine[J]. Journal of Nutrition,1995,( 5): 1238 1244.
[57] KELLY C P,CHETHAM S,KEATES S,et al. Survival of anti clostridium difficile bovine immunoglobulin concentrate in the human gastrointestinal tract[J]. Antimicrobial Agents & Chemotherapy,1997, 41(2): 236-241.
[58] LEE J,KANG H E,WOO H J. Stability of orally administered immunoglobulin in the gastrointestinal tract[J]. Journal of Immunological Methods,2012,384(1-2): 143-147.
[59] 张春刚,王加启,刘光磊,等. 乳品加工技术对乳中免疫球蛋白的影 响[J]. 中国畜牧兽医,2007,( 9): 64-66.
[60] 唐蓉,李晓璐,梁智铭,等. 不同加工方式对驼乳营养及功能成分的影 响[J]. 中国乳品工业,2024,52(9): 42-49.
[61] DIDIER L,ANNIE L,Halima E,et al. Immunochemical quantification of heat denaturation of camel (Camelus dromedarius) whey proteins[J]. Journal of Dairy Research,2006,73(1): 1-9.
[62] WEBER A,ENGELMAIER A,HAINDL S. Long-term stability of immunoglobulin G antibodies against bacterial antigens in human immunoglobulin G-deficient serum[J]. Journal of Allergy and Clinical Immunology,2020,145(2): AB177.
[63] MA H,Ó’FáGáIN C,O’KENNEDY R. Antibody stability: a key to performance-analysis,influences and improvement[J]. Biochimie, 2020,177: 213-225.
[64] 刘伟. 牛初乳免疫球蛋白(IgG)稳定性的研究及改善[D]. 北京: 中 国农业大学,2005. 68
[65] 张和平,杜文,郭军,等. 免疫乳中IgG的稳定性及蔗糖的保护作 用[J]. 中国乳品工业,2002,30(6):3-7.
[66] WONG P H,WHITE K M. Impact of immunoglobulin therapy in pediatric disease: a review of immune mechanisms[J]. Clinical reviews in allergy & immunology,2016,51: 303-314.
[67] LONG Y,WANG C,XIAO J,et al. Case report: novel multi exon homozygous deletion of ZBTB24 causes immunodeficiency, centromeric instability,and facial anomalies syndrome 2[J]. Frontiers in Immunology,2025,16: 1517417.
[68] ZHOU X,SHAN N N. The intersection of epigenetics and immune thrombocytopenia: new insights into disease mechanisms and treatments[J]. Molecular biology reports,2025,52(1): 257.
[69] CHEN J,WANG Y,CHENG J,et al. Synergistic impact of macrolide resistance and H3N2 infection on M. pneumoniae outbreak in children[J]. Microbiology Spectrum,2025,13(4): 0184424.
 返回顶部
返回顶部
 关注我们
关注我们